


| name | Amanita lepiotoides |
| name status | nomen acceptum |
| author | Barla |
| english name | "Large-Scaled Amidella" |
| images |
 1. Amanita lepiotoides  2. Amanita lepiotoides  3. Amanita lepiotoides |
| cap | The cap of Amanita lepiotoides is 60 - 120 mm wide, hemispheric, then convex, dry, with a decurved margin, becoming striate at maturity The cap is pure white, then cream with a tint of pink tint or ochraceous, and fauve-brown or grayish fauve-brown. The flesh is thick, white at first, then reddish, salmon or red-brown on exposure to air or on bruising. The volva is present as large, thick floccose/felted scales over the cap's center and fine fibrillose scales near the margin. |
| gills | The gills are free, rather distant, cream or yellowish white or pale white, rather large, thick, and almond shaped. The short gills are sometimes uncommon, rounded to rounded truncate to squarely truncate, and numerous. |
| stem | The stem is 100 - 200 × 10 - 25 mm, slender, subcylindric, and yellowish white to concolorous with the cap. The flesh is white turning salmon and stuffed to hollow. The ample, thick, membranous, subglobose volva often has an inner layer of floccose/felted material. |
| odor/taste | The odor of this species is weak or absent. The taste is said to be disagreeable. |
| spores | The spores measure (7.3-) 8.3 - 13.8 (-21.0) × (4.8-) 5.5 - 7.7 (-9.1) µm and are ellipsoid to elongate (occasionally cylindrical, occasionally broadly ellipsoid) and amyloid. Clamps are uncommon at bases of basidia. |
| discussion |
Amanita lepiotoides was originally described from mountains in southern France (Dép. Alpes-Maritimes). It is known from other localities in southern France as far west as the Pyrénées Orientales and from Italy. It is reportedly associated with Pine, Fir, Beech, and (possibly) Chestnut. This species is notable for the thickness of the inner layer of the volval, fragments of which are often left on the cap. Along with A. curtipes E.-J. Gilbert, A. ponderosa Malençon & R. Heim in Malençon, and A. valens (E.-J. Gilbert) Bertault of the Mediterranean region, A. lepiotoides is more closely related to the type (A. volvata (Peck) Lloyd) of section Amidella than is Amanita ovoidea (Bull. : Fr.) Link, another European species currently believed to belong in this section.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita lepiotoides | ||||||||
| author | Barla. 1885 Bull. Soc. Mycol. France 1: 193-4. | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Large-Scaled Amidella" | ||||||||
| synonyms |
≡Amanitopsis lepiotoides (Barla) Sacc. 1887. Syll. Fung. 5: ??.
≡Pseudofarinaceus lepiotoides (Barla) Kuntze. 1891. Rev. Gen. Plant. 2: 868.
≡Vaginata lepiotoides (Barla) Kuntze. 1898. Rev. Gen. Plant. 3(2): 539.
≡Amidella lepiotoides (Barla) E.-J. Gilbert. 1940. Iconogr. Mycol. (Milan) 27, suppl. (1): 77, tab. 23 (fig. 6).
?=Squamanita cettoiana M. M. Moser nom. prov. 1978. Kl. Kryptog. 4. Aufl. 2 (b/2): 251. The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||
| MycoBank nos. | 155691, 178535, 284135 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes |
in herb. Boudier => PC??, not found by recent investigators, lost? [Note: topotype, S; other authentic material in BPI, NICE per Neville and Poumarat (2004), NY.] | ||||||||
| selected illustrations |
Barla. 1888. Fl. Mycol. Illus. Champ. Alpes-Marit. 1: Pl. 8 bis. Galli. 2000. Boll. Gruppo Micol. G. Bresadola 43(2): 102. Zecchin. 2000. Boll. Gruppo Micol. G. Bresadola 43(2): 166, 167. | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material is derived from the protolog of the present species, (Gilbert 1940 & 1941), and original research of R. E. Tulloss. The source for the latter is an unpublished Ms. developed with guidance from Dr. Bas between 1985 and the early 1990's; it is still in unfinished form. The reader will note that when it was drafted, RET had not yet developed some relevant methodology and had not yet adopted the associated notation. RET decided to post this information with comments because it is unknown whether he will ever have time to commit to resuscitating in full scale his project on sect. Amidella. [Note: At some point after the Ms. was well under way, I became concerned about whether I had accurately segregated vascular hyphae from septate hyphae that happened of have yellowish refractive walls. At that point, I substituted the phrase "refractive hyphae" for "vascular hyphae" in my older Mss. in contexts in which I thought I might have misapplied the latter term.—RET] | ||||||||
| pileus | 60 - 120 mm wide, hemispheric, then convex, pure white then cream with a pink tint to ochraceous and, finally, fauve-brown or grayish fauve-brown, dry; context thick, fleshy, white at first, then reddish, salmon, or red-brown on exposure to air or on bruising; margin decurved, sterile for a short space, appendiculate with white flocculence, nonstriate or slightly excoriated at first, becoming striate at maturity; universal veil as large (20 mm or more across), rather thick, floccose/felted scales over disc reduced to fine fibrillose scales near margin, white at first, then becoming brownish or reddish brown (taking on the appearance of Lepiota procera according to some authors in case volva entirely covers the pileus and outer layer of the volva ruptures), rarely as patch of outer layer of universal veil superimposed on scales of inner layer material. | ||||||||
| lamellae | free, rather distant, cream or yellowish white or pale white like beeswax, bruising rather quickly to pinkish fauve then becoming brownish purple, drying brown (e.g., 6D4 or 6E4-5 or slightly grayer to 6E7) to very dark brown to blackish, rather large, thick (up to 2mm near stipe), almond shaped, often forking, margin flocculose with white material; lamellulae sometimes uncommon, rounded to rounded truncate to squarely truncate, numerous, often anastomosing to lamellae. | ||||||||
| stipe | 100 - 200 × 10 - 25 mm, slender, subcylindric, flaring at apex, yellowish white to concolorous with pileus, finely flocculose above, flocculose/scaly below, decoration concolorous with universal veil remnants on pileus, with surface sometimes cracking into recurved scales; context white turning salmon or reddish or reddish brown on bruising, stuffed to hollow; partial veil fugacious, as ring of white particles in young basidiocarps or somewhat thicker and bearing striations on upper surface, quickly collapsing and disintegrating; universal veil as ample, thick, brownish, membranous, subglobose volva often with inner layer of floccose/felted material rather thick and separating, up to 49 mm or more wide, top point of limb up to 73 mm or more from base of stipe. | ||||||||
| odor/taste | Odor absent or weak. Taste disagreeable. | ||||||||
| macrochemical tests |
NH4OH - on pileipellis and pileus context slowly becoming an intense, deep red (like red ink), then becoming brownish. | ||||||||
| pileipellis | filamentous undifferentiated hyphae 3.5 - 7.9 µm wide, subradially oriented, gelatinizing, sometimes yellowish brown; refractive hyphae up to 3.5 µm wide. [Note: This description predates my current understanding of the structure of the pileipellis in Amanita.—RET] | ||||||||
| pileus context | branching undifferentiated filamentous hyphae 1.8 - 7.0 µm wide; acrophysalides ovoid to broadly ellipsoid, up to 78 × 60 µm; refractive hyphae plentiful. | ||||||||
| lamella trama | bilateral, divergent; broad, divided into three roughly equal zones, with central stratum comprising interwoven hyphae and narrowly inflated ?terminal cells; divergence restricted to two lateral zones, as smooth even curve ending perpendicular to hymenial surface; undifferentiated filamentous hyphae 1.5 - 12.5 µm wide, branching, sometimes with rather short segments, sometimes with walls thickened slightly (to about 0.5 µm); inflated cells as slightly inflated intercalary hyphal segments or as terminal clavate to narrowly clavate to subventricose to ventricose to ventricose-rostrate to occasionally ellipsoid cells, up to 139 × 28 µm, thin-walled or sometimes with walls up to 0.8 µm thick, often curved and part of smoothly curving divergence of trama; refractive hyphae 1.2 - 8.7 µm wide, branching, sometimes plentiful. | ||||||||
| subhymenium | as sporulation begins, ramose, comprising cylindrical hyphal segments (4.0 - 6.5 µm wide) in branching chains roughly perpendicular to the central stratum; in more mature material with some basidia arising from partially inflated or small ovoid to subcoralloid inflated cells, with previously noted branching structure still evident; refractive hyphae branching, sometimes rather plentiful. | ||||||||
| basidia | 37 - 58 × 7.2 - 18.9 µm, usually 10.5 - 12.5 µm wide, near apex, 4-, or occasionally 3- or 2-sterigmate, thin-walled, rather narrow below, at times with sudden expansion near apex, some with refractive contents, some containing brown or orange-brown or yellow-brown or dark brown necropigment at times as spherule or spherules; sometimes surrounded with interwoven, refractive hyphae that completely penetrate the hymenium; clamps rare. [Note: It now seems to me, especially considering the age of the material that I was examining, that the above-mentioned refractive hyphae could have been invasive elements of a mold that had attacked the exsiccatum.—RET] | ||||||||
| universal veil | On stipe base, exterior layer: easily separating on reinflation in KOH, comprising filamentous undifferentiated hyphae 4.4 - 13.1 µm wide, longitudinally arranged, gelatinizing, with some slightly inflated; inflated cells rare or absent. On stipe base, interior layer: undifferentiated filamentous hyphae 1.8 - 20.1 µm wide, dominating, branching, interwoven; inflated cells ellipsoid to subglobose, up to 93 × 88 µm; refractive hyphae 1.8 - 18.4 µm wide. On stipe base, inner surface layer: filamentous undifferentiated hyphae 5 - 15 µm wide, sublongitudinally oriented, somewhat interwoven, brownish; inflated cells longitudinally oriented, scattered, subcylindric to subfusiform. On pileus, outer layer: similar to interior layer on stipe base with upper surface somewhat gelatinizing and inflated cells yellow-brown and more elongate (up to 200 × 40 µm). On pileus, inner surface layer: filamentous undifferentiated hyphae 1.5 - 4.0 µm wide, slightly gelatinizing brownish-yellowish, somewhat interwoven, subradially arranged. | ||||||||
| stipe context | longitudinally acrophysalidic; filamentous, undifferentiated hyphae 2.0 - 14.0 µm wide, frequently branching; acrophysalides rather short, to 125 × 43 µm, clavate to ovoid, thin-walled or infrequently with walls up to 1.0 µm thick; refractive hyphae 3.5 - 14.5 µm wide, branching, plentiful. | ||||||||
| partial veil | filamentous undifferentiated hyphae 3.5 - 12.2 µm wide, frequently branching, tangled; inflated cells narrow ellipsoid to ovoid to globose, to 70 × 63 µm; refractive hyphae 5.2 - 7.0 µm wide, locally in dense tangles. | ||||||||
| lamella edge tissue | not described. | ||||||||
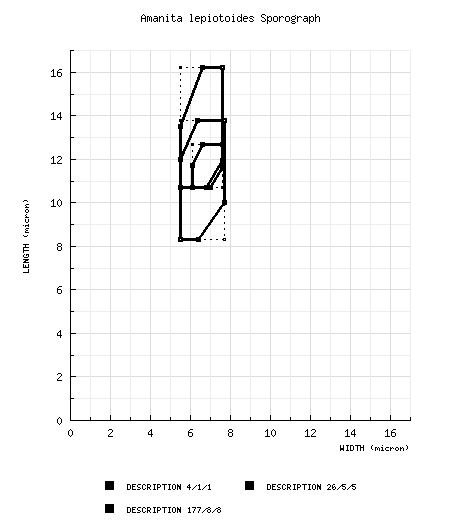
| basidiospores |
Gilbert (1940) spores from lectotype: [4/1/1] 10.7 - 12.7 × 6.1 - 7.6 μm, (L = 11.7 μm; W = 6.9 μm; Q = 1.53 - 1.92; Q = 1.70). [Note: Data derived by measuring the spores drawn in lateral view from tab. XXIII (fig. 6).—ed.] Gilbert (1940): [26/5/5] 10.7 - 16.2 (-16.6) × (4.6-) 5.5 - 7.6 μm, (L = 11.6 - 15.1 μm; L' = 12.7 μm; W = 5.7 - 7.5 μm; W' = 6.4 μm; Q = (1.53-) 1.57 - 2.46 (-2.47); Q = 1.70 - 2.28; Q' = 2.0). [Note: Data derived by measuring the spores drawn in lateral view from tab. XXIII (fig. 6) and tab. XXIV (figs. 1-5). Spores from A. lepiotoides f. macrospora E.-J. Gilbert nom. inval. are not included here.—ed.] Neville and Poumarat (1940 & 1941), for some reason excluding at least some data on 260 spores which are from material in NICE treated in a separate table on p. 668: [170/-/-/-] (8.5-) 9 - 12.5 (-13) × 6 - 8 μm, (L' = 10.6 μm; W' = 6.8 μm; Q = (1.30-) 1.36 - 1.80 (-2.0); Q = 1.48 - 1.61; Q' = 1.55). RET: [177/8/8] (7.3-) 8.3 - 13.8 (-21.0) × (4.8-) 5.5 - 7.7 (-9.1) µm, (L = (9.2-) 9.7 - 13.0 µm; L’ = 10.8 µm; W = 6.0 - 6.9 (-7.0) µm; W’ = 6.4 µm; Q = (1.18-) 1.30 - 2.18 (-3.55); Q = (1.40-) 1.43 - 2.05; Q’ = 1.69), amyloid, sometimes dextrinoid when immature, sometimes containing dark necropigment when damaged, smooth, thin-walled, hyaline to pale dingy yellow, ellipsoid to elongate, occasionally cylindrical, occasionally broadly ellipsoid, sometimes with a hook at the apex terminating in the apiculus, sometimes swollen at one end; apiculus sublateral, cylindrical; contents granular, sometimes golden-guttulate in Melzer’s Reagent; white in deposit. | ||||||||
| ecology |
Apparently rather rare, although Gilbert (1941: 288) reports an occasion on which a troop of ca. 20 basidiomes were seen by the collector A. Thouvenin. In June to September and November (Assoc. Myc. Faç. Méd., 1986), solitary to subgregarious. France: At 700 - 750 m elev. On silicaceous, acid soil associated with Pinus and Castanea. Italy: At edge of mixed woods (Fagus, Picea excelsa, and Pinus sylvestris) on acid soil near meadow. In association with Quercus pedunculatus (Parrot, 1960) and on clayey-calcareous soils in association with Quercus, Carpinus and Castanea (Massart, 1984) in southwestern France. On Col de l’Ouillat, Pyrenées Orientales, France under Quercus and Fagus (Marchand, 1973). The packet containing Marchand’s 1968 collection from Les Albères (north face of the col) includes notes indicating that another was found the previous day in acid humus in a Fagus forest with some Quercus nearby. Bertault (1964) says he has never found A. lepiotoides in Morocco. Gilbert (1940 & 1941: 287-288) provides an extensive discussion of materials (collections, watercolors, book illustrations) that he was able to locate. Collections identified as in Gilbert's personal collection must be assumed to be lost with his herbarium, notebooks, etc. Among other collections Gilbert mentions, material in Barla's herbarium (NICE) was also found by Neville and Poumarat (2004: 662). | ||||||||
| material examined |
Gilbert (1940: 126-129): FRANCE: ALPES MARITIMES—Berre-les-Alpes [ca. 700 m], 21.vi.1881 J.-B. J. J. Barla t. 70 (lectotype, in herb. Boudier => ?PC, ?lost), s.d. J.-B. J. J. Barla "n. 2?" (in herb. Barla? => NICE), 19.vii.1930 E.-J. Gilbert s.n. (in herb. Gilbert, exsiccatum and basidiome, lost).
DOUBS—Besançon, RET: FRANCE: ALPES MARITIMES—Berre les Alpes, | ||||||||
| discussion |
The difficulty in getting a good understanding of this species is complicated by its apparent rarity and the fact that the majority of the collections we have found in herbaria are poorly preserved or include only immature or overly mature specimens. The impact is particularly severe on the spore data which probably misrepresents the average spore size and shape by making it appear that spores are shorter and less elongate than they would be in well-dried, well-preserved collections. The material from Lignan-de-Bordeaux was poorly preserved; many of the spores were broken. Only thirteen complete spores were found on the material. The Barla specimen in NY is at the stage in which sporulation was just beginning; again the spore measurements are probably skewed to the low side. The 1978 Cetto collection and the Barla collection in BPI are incompletely expanded “buttons.” The topotype of 1895 appears to be immature. The 1888 Barla collection from Nice was annotated “juvenilis.” The 1979 Cetto collection was over-mature and had its hymenium damaged by insects after drying. The Albanesi collection also has rather small spores, possibly due to its infection by a mold (reported in a note of J. Trimbach included in the herbarium packet). The exceptions are the collections of Huijsman and Marchand and the Barla collection of 1887 from Nice which yielded the highest values measured for L (11.5 µm, 13.0 µm, and 12.0 µm respectively) and Q (1.90, 2.05, and 1.75 respectively). more | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita lepiotoides |
| bottom links |
[ Section Amidella page. ] [ Amanita Studies home. ] [ Keys & Checklists ] |
| name | Amanita lepiotoides |
| bottom links |
[ Section Amidella page. ] [ Amanita Studies home. ] [ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.